For the most part sharks have laterally positioned eyes, ie: the
eyes are positioned on the sides of the head. However, some of the
more benthic species (eg: orectolobids and squatinids) have more
dorsally positioned eyes.
When compared with mammals, sharks are considered to have
generally small eyes compared to body size, although some species of
Lamniforme shark have much larger eyes, including the great white
and Mako sharks. In all
sharks the two eyes oppose each other which allows for 360◦ visual
field, especially in the case of a shark in motion utilising a
laterally sinusoidal swimming pattern.
Limited eye movements are observed in most species, primarily
to compensate for the swimming movements and stabilise the visual
field (Harris, 1965).
Binocular overlap is small and blind areas exist just in front of
the snout and behind the head when the animal is still.
The size of these blind areas depends on the shape of the
head and the configuration of the eyes, but typically the forward
blind area extends less than one body length a head of the rostrum
(Carrier et al., 2004).
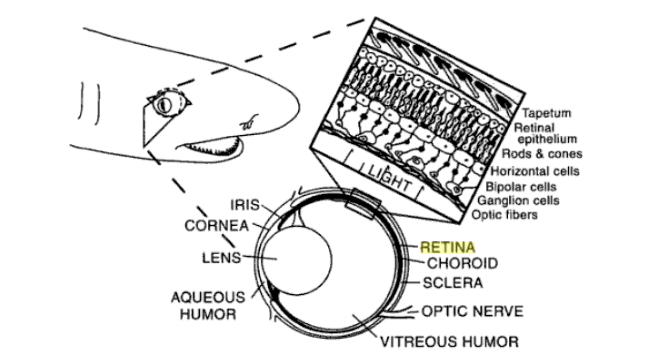
Fig 1: Cross section through a shark eye showing ocular and retinal
anatomy. Tapetum lucidium shown in non-occluded state exposing
reflective plates for greater visual sensitivity under scotopic
conditions. (Heuter and Gilbert, 1990)
Visual Performance
 Sharks
cannot actually see in colour but instead use contrast in colour to
assess their surroundings (Hart et al., 2012).
Certainly the importance of vision in the daily lives of
sharks finds support in the anatomical and physiological visual
adaptations, many of which appear to be correlated with species
behaviour and ecology.
Sharks
cannot actually see in colour but instead use contrast in colour to
assess their surroundings (Hart et al., 2012).
Certainly the importance of vision in the daily lives of
sharks finds support in the anatomical and physiological visual
adaptations, many of which appear to be correlated with species
behaviour and ecology.
Oceanic and deep-sea sharks have the largest
eyes amongst sharks and presumably rely more heavily on vision than
coastal and benthic species, while variation in the ratio of rod and
cone photoreceptors and the spatial resolving power of the eye all
appear to be closely related to differences in habitat and
lifestyle.
There is evidence that ontogenetic changes in the visual system,
such as changes in the spectral transmission properties of the lens,
lens shape, focal ratio, visual pigments and spatial resolving
power, allow elasmobranchs to adapt to environmental changes imposed
by habitat shifts and niche expansion.
Anatomy of a Shark Eye
The outer layer of the shark eye comprises a thick cartilaginous
sclera and a gently curving, transparent cornea, the fine structure
of which includes sutural fibers that resist corneal swelling and
loss of transparency in challenging chemical environments (Tolpin et
al. 1969).
Pupil
Unlike teleosts (bony fish), most sharks have a dynamic iris that
can increase the size of the pupil in dim light or decrease it in
bright light. The shape
of the pupil varies amongst species depending upon their respective
feeding strategies. The
pupil can be circular (eg. Most deep sea sharks, which have less
mobile pupils for more constant, low light conditions), vertical
slit (eg. Carcharhinus spp., Negaprion brevirostris), horizontal
slit (eg. Sphyrna tiburo), oblique slit (eg. Schyliorhinus canicula,
Glinglymostoma cirratum), or crescent-shaped (eg. Many skates and
rays) (Carrier et al., 2004).
Mobile slit pupils are typically found in active predators
with periods of activity in both photopic (bright light) and
scotopic (dim light) conditions, such as the Lemon shark, N.
brevirostris (Gruber, 1967); a slit pupil that can be closed down to
a pinhole is thought to be the best way to achieve the smallest
aperture under photopic conditions, because a circular pupil is
mechanically constrained from closing to a complete pinhole (Walls,
1942).
Eyelids (Ocular Adnex)
The ocular adnexa (eyelids and adjacent structures to the eye) are
well developed in sharks, however the upper and lower eyelids do not
move appreciably to cover the entire eye (Gilbert, 1963).
Benthic species like the wobbegongs (Orectolobids) have more
mobile lids, which serve to protect the eyes whilst burrowing.
Some shark species, especially the carcharhinids and
sphyrinids, posses a third eyelid, the nictating membrane, which can
be extended from the lower nasal corner of the eye to cover the
exposed portion of the eye (Gilbert, 1963).
The nictating membrane acts to protect the eye during feeding
and mating.
Cornea
The shark cornea is virtually absent underwater due to its
similarity in refractive index to the seawater (Heuter, 1991),
leaving the crystalline lens to provide the total refractive power
of the eye. Shark
lenses are typically large, relatively free of optical aberration,
and ellipsoidal in shape, although the spiny dogfish, Squalus
acanthias, and clearnose skate, Raja eglanteria, have nearly
spherical lenses (Sivak, 1991).
Some shark lenses contain yellowish pigments that are
enzymatically formed oxidation products of tryptophan, similar to
lens pigments found in many teleosts and diurnal terrestrial
animals. These pigments
filter near ultraviolet light, which helps to minimise defocus of
multiple wavelengths (Chromatic aberration), enhance contrast
sensitivity, and reduce light scatter and glare under conditions of
bright sunlight (Zigman, 1991).
They may also help to protect the retina from UV damage in
shallow benthic or epipelagic species.
Choroid
At the back of the shark eye behind the retina and in front of the
sclera lies the choroid, the only vascularised tissue within the
adult shark eye. The
shark retina is not vascularised and typically contains no landmarks
other than the optic disk (corresponding to a small blind spot in
the visual field), which contains no photoreceptors and marks the
exit of retinal ganglion cell fibres via the optic nerve from the
retina to the CNS. The
choroid in nearly all sharks contains a specialised reflective layer
known as the tapetum lucidium, which consists of a series of
parallel, platelike cells containing guanine crystals (Gilbert,
1963). The function of
the tapetum lucidium is to reflect back those photons that have
passed through the retina and have not been absorbed by the
photoreceptor layer, allowing a second chance for the detection of
photons and thereby increasing the sensitivity of the eye in dim
light.
Retina and the CNS (Central Nervous System)
 The largest impact on our understanding of visual capabilities in
sharks came with the eventual finding that practically all sharks
have duplex retinas containing both rod and cone photoreceptors
(Gruber and Cohen, 1978), beginning with the unequivocal evidence of
cones in the lemon shark (N. brevirostris) retina presented by
Gruber et al. 1963.
Cones subserve photopic and colour vision and are responsible for
higher visual acuity; rods subserve scotopic vision and are involved
in setting the limits of visual sensitivity in the eye.
Prior to Gruber’s work in 1963, sharks were thought to
possess all rod retinas, and thus were considered to have poor
visual acuity and no capability for colour vision, which we now know
to be untrue.
The largest impact on our understanding of visual capabilities in
sharks came with the eventual finding that practically all sharks
have duplex retinas containing both rod and cone photoreceptors
(Gruber and Cohen, 1978), beginning with the unequivocal evidence of
cones in the lemon shark (N. brevirostris) retina presented by
Gruber et al. 1963.
Cones subserve photopic and colour vision and are responsible for
higher visual acuity; rods subserve scotopic vision and are involved
in setting the limits of visual sensitivity in the eye.
Prior to Gruber’s work in 1963, sharks were thought to
possess all rod retinas, and thus were considered to have poor
visual acuity and no capability for colour vision, which we now know
to be untrue.
Spatial Topography of the Retina
The spatial topography of retinal cells can, reveal much about the
quality of vision in these animals.
Although sharks do not have all cone foveas, they do have
retinal areas of higher cone and/or ganglion cell density, which
indicates regional specialisations for higher visual acuity (Collin,
1999). Higher cone
concentrations have been found in the central retina of the nurse
shark (G. cirratum) (Gruber, 1965),
white spotted bamboo shark (Chiloscyllium plagiosum) (Yew et al.
1984) and the white shark (Carcharodon carcharhias) (Gruber and
Cohen, 1985). Franz
(1931) was the first to report horizontal streaks of higher ganglion
cell density in the small spotted catshark (Scyliorhinus canicula)
and the smooth hound (Mustelus mustelus).
The horizontal visual streak is an adaptation for more or
less two dimensional terrain environments such as the sea bed or the
sea surface. Concentric
retinal areas are more applicable for imaging a limited spot in the
visual field or for operating in complex, three dimensional visual
environments, such as reefs or the open water.
Both, the cookie cutter shark and the white shark are ambush
predators in open water, and both appear to have retinal areas, not
streaks (Carrier et al., 2004).
Accommodation
Accommodation
is the process by which the vertebrate eye changes optical power to
maintain a clear image (focus) on an object as its distance changes.
Accommodation acts like a reflex, but can also be consciously
controlled, in humans at least. Mammals, birds and reptiles vary the
optical power by changing the form of the elastic lens using the
ciliary body. Sharks
that accommodate do not change the shape of the lens, but instead
change the position of the lens by moving it toward the retina (for
distant targets) or away from the retina (for near targets).
The lens is supported dorsally by suspensory ligaments and
ventrally by the pseudocampanule, a papilla with ostensibly
contactile function (Sivak and Gilbert, 1976).
